Получивший широкой резонанс в среде профильных профессиональных популяционных генетиков и простых любителей препринт статьи Lazaridis et al. 2013 содержит огромное количество важных наблюдений и заключений насчет происхождения и эволюции структуры генофонда современных европейцев. Объективности ради стоит отметить, что наверное не менее половины выводов вышеупомянутой статьи были озвучены ранее персональными геномными блоггерами в ходе более ранних самостоятельных экспериментов и анализов древней ДНК. Но важность статьи Lazaridis et al. 2013, конечно же, в другом. В ней приводятся новые данные ( результаты изучения древнего ДНК) ряда доисторических популяций групп людей Европы. В частности, были приведены результаты изучения древнего ДНК мезолитических европейских охотников-собирателей из Люксембурга, чьи мтДНК были опубликована несколько лет назад; результаты генотипирования неолитического образца ДНК неолитического земледельца из Германии: наконец, препринт статьи содержал данные сразу по нескольким мезолитическим охотникам-собирателей из Швеции.
По раскладу своих предковых компонентов, люксембургский образец похож на образцы La Brana (иберийский палеолит). В свою очередь, ДНК шведов периода мезолита похожа на ДНК шведских неолитических охотников-собирателей. Аналогично, структура предковых компонентов у неолитического земледельца напоминает распределение компонентов у тирольского ледового человека Этци, шведского неолитического земледельца культуры воронковидных кубков и современного населения Сардинии. Недавно опубликованный верхнепалеолитический образец жителя стоянки Мальта (Прибайкалья), авторы относят к отдельному генетическому супер-компоненту, который в статье именуется «компонентом древнего северо-евразийского населения», Согласно наиболее вероятной из обсуждаемых в статье моделей, носители этого компонента смешались с западными евразийцами еще до того, как носители мезолитического европейского компонента смешались с неолитическими земледельцами с ближнего Востока.
Как я отмечал ранее, очевидность результатов вряд ли нуждается в отдельных комментариях. Положение индивидов по степени их сходства (выраженного посредством параметра z D-статистики) относительно условной оси на одном конце которых находятся южные европейцы, на другом — северные европейцы. Шведские сэмплы древнего ДНК времен мезолита и пост-мезолитической (переходной к неолиту) культуры ямочной керамики Готланда сдвигаются в сторону северных европейцев (представлены референсной популяцией литовцев). Примечательно, что в ту же стороны сдвигаются и представители иберийско-испанского мезолита (образцы La Brana 1 и La Brana 2).
Образцы древнего ДНК представителей культуры воронковидных кубков, Эци Тирольца закономерно смещаются к другому полюса спектра — южным европейцам (которые представлены сардинцами).
PCA график дает отличное представление о соотношении различных компонентов:

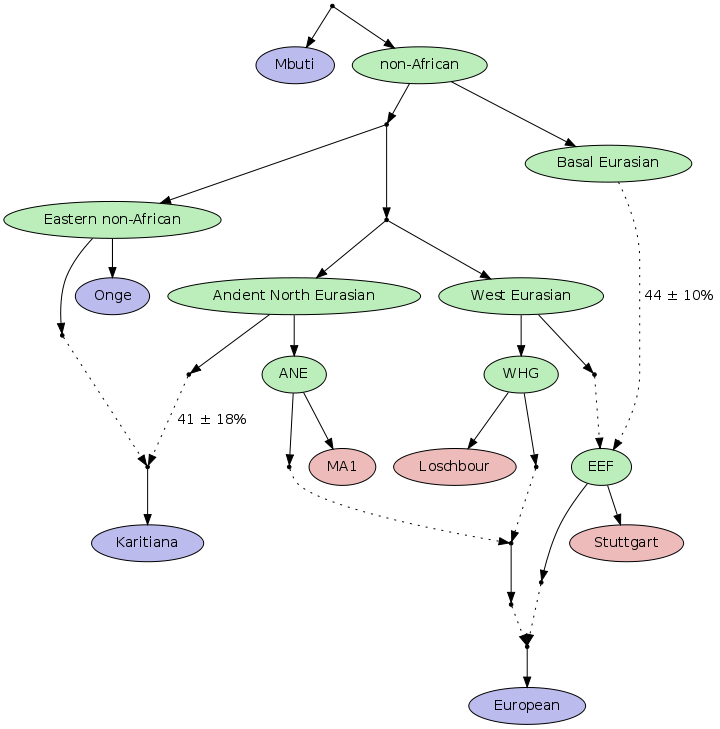
Таким образом, костяк европейского генофонда образован за счет сочетания трех компонентов:
мезолитические охотники-собирателей Европы (WHG) + древние северо-евразийские популяции между Уралом, Центральной Азией и Сибирью (AHE) + неолитический компонент (генетически связанный с ближневосточными земледельцами EEF).
Поскольку в нашем случае мы имеем три исходных компонента, то любая европейская популяция может быть представлена в виде тримодального распределения этих компонентов. Визуализация этого распределения достигается путем отображения популяций внутри треугольника (каждый из углов которого представляет собой отдельный чистый предковый компонент). Таким образом мы можем отображать не только группы людей (т.е популяции), но и отдельных современных индивидов с генотипированными снипами. При визуальном изучении расположения популяций внутри треугольника, мы можем отметить cмещение спектра разнообразия в сторону предкового компонента неолитических земледельцев (EEF). Это наблюдение еще раз подтверждает насколько важным событием для эволюции и развития европейского генофонда являлась неолитическая революция. Она принесла с собой не только технологические изменения, но и перемены в генофонде тогдашних европейцев. Однако если быть более точным, то нет никаких сомнений в том что компонент EEF аккумулирует в себя не только генофонд первых европейских земледельцев, но и остаточный момент от смешения этого компонента с представителями четвертого компонента («базальных евразийцев»).

Выявленные предковые компоненты отличаются значительной дискретностью, и в своем чистом виде практически не перекрываются. Именно по этой причине именно эти компоненты (а не общепринятые ныне в аутосомных исследованиях попгенетиков этногеографические компоненты) могут использоваться в качестве неколлинеарных факторов в анализе эволюции генофонда отдельных народов.
- Компонент западноевропейских охотников-собирателей мезолита ( WHG ): метапопуляция этого компонента включает в себя образец Loschbour (мезолитический Люксембург, 8000 лет до настоящего времени) и два мезолитических образца древнего ДНК людей из пещеры La Brana в Испании. Тем не менее, в настоящий момент своего пика WHG (почти 50%) достигает среди эстонцев и литовцев , на востоке Балтийского региона. В этом смысле эти популяции являются наследниками древнейших жителей мезолита Европы. К этой группе примыкает группа шведских неолитических популяций (скандинавские охотники-собиратели (SHG ) : эта мета- субпопуляция состоиь из шведских мезолитических и неолитических образцов ДНК из Моталы и Готланда , соответственно. Судя по всему, здесь мы имеем дело с более восточным вариантом WHG , с небольшой примесью генов от древних северо-евразийских популяций.
Удельная доля компонента WHG в генофонде популяций по мере убывания (градиент убывания в направлении с северо-запада на юго-восток Европы).
Эстонцы 0,495
Литовцы 0,464
Исландцы 0.456
Беларусы 0,431
Норвежцы 0,428
Испанцы 0,068
Греки 0,058
Мальтийцы 0
Ашкенази 0
Сицилийцы 0
- Компонент ранних европейских земледельцев (EEF) : по всей видимости, это гибридный компонент являющийся результатом смешивания части загадочных «базальных евразийцев» и носителей компонента WHG где-то в Европе, возможно, на Балканах. Метапопуляция EEF в чистом виде представлена у представителя линейно-ленточной керамики (Штутгарт, Германия), Этци Тирольского человека, и неолитического земледельца культуры воронковидных кубков. В наше время пик это компонента приходится на Сардинию, Сицилийцев, ашкеназов и жителей Мальты (примерно 80-90%).
Удельная доля компонента EEF в генофонде популяций по мере убывания (градиент убывания частоты направлен в сторону обратную WHG, т.е с юго-востока Европы на северо-запад, однако градиент выражен гораздо менее четко).
Мальтийцы 0,932
Ашкеназим 0,931
Сицилийцы 0,903
Сардинцы 0,817
Испанцы 0,809
Норвежцы 0,411
Исландцы 0,394
Шотландцы 0,39
Литовцы 0,364
Эстонцы 0,322
3. Компонент древних северо-евразийцев ( ANE ): метапопуляция компонента включает в себя 24000 летний верхне-палеолитический образец охотника-собирателя из южно-центральной Сибири, принадлежащий к Y- ДНК гаплогруппе R *, (MA -1), а также верхнепалеолитический образец из центральной Сибири (Афонтова Гора -2) ( AG2 ). Этот компонент мог вероятно присутствовать в Южной Скандинавии по крайней мере со времен мезолита, но Западной Европы достиг уже в конце эпохи неолита. В современной Европе самый высокий процент это компонента наблюдается у эстонцев (на уровне чуть более 18%), и достигает такого же уровня среди шотландцев.
Эстонцы 0,183
Шотландцы 0,182
Венгры 0,179
Литовцы 0,172
Чехи 0,167
Итальянцы из Бергамо 0,108
Сицилийцы 0,097
Ашкеназим 0,069
Мальтийцы 0,068
Сардинцы 0,008
Сводная таблица по раскладу компонент у отдельных популяций и их формальной статистической значимости (f3-статистика):

Эпилог
В апреле и мае 2012 года (задолго до появления статьи Lazaridis et al. 2013), в ходе изучения структуры кластеров компонентов в древних ДНК (чьи SNP-данные были тогда у меня в наличии). В ходе анализа Admixture K=3 программа дала интересное распределение по современным популяциям и древним образцам. Как видно из географического распространения, кластер-метопапуляции готландских охотников-собирателей/ мезолита и современных саамов в значительной части перекрывается с ареалом современного пика компонентов мезолитических охотников-собирателей. Неолитический (средиземноморский) компонент совпадает с ареалом компонента EEF. Наконец, контуры ареал компонента который я ошибочно обозначил как Corded Ware Indo-European, практически перекрываются контурами ареала ANE (пик которого, как отмечалось выше, приходится на популяции шотландцев и эстонцев).


.

Для отправки комментария необходимо войти на сайт.