Ув. Вячеслав Носевич! Вы высказали обоснованную критику моих взглядов на происхождение динарского кластера I2a. Считаю нужным высказать свои встречные возражения. Мои соображения основаны не на голых спекуляциях, а строгих математических формальных методах. Собственно именно наиболее вероятная, парсимоническая трактовка полученных результатов филогенетического и статистического анализа молекулярного разнообразия гаплотипов I2a2 свидетельствует о экспансии этой субклады с территории Карпато-полесского региона. Во-первых, нужно сразу оговорится что коль речь идет о славянах из Восточной Европы, то речь идет не просто о I2a, а о I2a2 — поскольку эта группа практически экслюзивна представлена у славян и их ассимилированных потомков. Далее — о статистическом анализе Остался мой вопрос незамеченым. Так вот, сравнение результатов теста AMOVA по 2 группировкам -лингвистической (популяции объединены в группы по принадлежности к той или иной языковой группе) и антропологической (популяции объединены по принципу генетической близости антропологических признаков)- показывает, что вероятность корреляции разнообразия отдельных этногрупп I2a2 с лингвистическими барьерами выше, чем с антропологическими. Хотя в обоих группировках 98% разнообразия приходится на вариации внутри популяций, однако при анализе статистической значимости вероятность верности нулевой гипотезы (о наличии вариации внутри популяций) равна только 0.005 (0.5%). Поэтому, с точки зрения статистики случайных чисел, следует признать этот результат статистически незначимым, т.е случайным. В процент разнообразия между этногруппами популяции выше именно в лингвистической группировке примерно в два раза -0.20% против 0.10% в антропологической группировке. При этом, в антропологической вероятность верности нулевой гипотезы, т.е. того, что группы этнопопуляций являются разнообразными в плане полиморфизмов Y-STR гаплотипов I2a2a, составляет примерно 14%. В то же время, как вероятность верности нулевой гипотезы о существенном разнообразии Y-STR гаплотипов I2a2a между группами популяций, разбитых по лингвистической близости, примерно в два раза выше и составляет 28%. Следуя полученным стат.результатам, необходимо признать, что I2a2a не являются автохтонами Балкан и Динарских Альп, в противном случае наблюдалась более существенная корреляция между популяцией I2a2a и антротипом. Дуализм антропологических параметров популяций Динарских Альп и языковой принадлежности — хорошо известный исследователям факт. Поскольку популяции I2a2a лучше коррелируют с языком, а не с антротипом, то можно сделать два вывода: 1) экспансия субклада произошла недавно, т.к. не утерялась связь представителей генетической линии с языком 2) поскольку славянские языки были явно привнесены на Балканы, то нужно признать, что I2a2a были в числе генетических линий, представители которых привнесли славянские языки на Балканы. Я проанализировал молекулярное и стандартное разнообразия, а также генетическую дистанцию гаплотипов представителей субклада I2a2-Dinaric. В ходе анализа мною проанализировано 624 «коммерческих» гаплотипа (17-67 маркеров) этого субклада (плюс некоторое количество гаплотипов из научных выборок), разбитых на этнопулы (согласно задекларированной национальности носителя гаплотипа). Комплексный анализ трех параметров позволяет, наряду с филогенетическим анализом, определить место вероятное происхождения субклада, а также предположить характер и степень влияния популяционных эфектов. Вот, к примеру, еврейский кластер с максимальной дистанцией и самым низким разнообразием указывает на недавнее происхождение кластера как следствие чистого эфекта отца-основателя. Боснийско-герцеговинский кластер (второй по величине интерпопуляционной дистанции), но с более высоким уровнем разнообразия указывает на более удаленный эфект основателся популяции. В то время, как те же хорватский кластерI2a2-Din с низким уровнем разнообразия, но с незначительной дистанцией от других популяций -следствие кумулятивного результата пресловутых популяционных эфектов основателя и бутылочного горлышка.
Ответ В.Носевича:
«Вадим, спасибо за ссылки! Это обсуждение я раньше не видел. Теперь Ваша позиция мне ясна, и я могу ее прокомментировать. Прежде всего хочу сказать, что отчасти Ваши претензии порождены недоразумением. Вам показалось, что я — сторонник происхождения динарского кластера из «ледникового убежища». Но перечитайте внимательнее мой текст — я этого вовсе не утверждаю, пишу только о неолитических культурах на местном мезолитическом субстрате. Почему мезолитическом? Да потому, что неолит анатолийского происхождения не мог содержать I2a. Уже установлено, что его представители имели G2a и E1b-V13, продолжают дискутироваться J2 и R1b, но никак не I2a. Чтобы избежать дальнейших недоразумений, и учитывая то, что Вы сделали эту дискуссию общедоступной, я постараюсь оговаривать даже очевидные вещи. Заранее прошу прощения, если что-то покажется тривиальным. Только не называйте меня профессором — я уж больше 20 лет простой кандидат наук, и им, наверное, умру. Во всяком случае, не испытываю потребности тратить время на защиту докторской или на доказывание кому-то, что достоин присвоения ученого звания. Итак, мы будем обсуждать происхождение так называемого динарского кластера гаплогруппы I2a. Раньше его выделяли по мутации M423, но сегодня уже известно, что эта ветвь делится на дочерние, из которых нас интересует L621/S392 и ее подветвь L147.2. Ее более поздним ответвлением, видимо, являются мутации L147.4 и L343. Произошли они до формирования динарского кластера или после — пока неясно, поскольку определены они у считанного числа индивидов. Но L147.2 выявлена, помимо славян, у как минимум одного англичанина (его Kit Number в соответствующем FTDNA-проекте — 14703), двух немев (162426 и 164573), одного итальянца (93943) и одного еврея из Литвы (211949), а L147.4 и L343 — только у славян: двух поляков (209633 и 76814) и боснийца (154978). При этом никого с L147.2+ на L147.4 и L343 не тестировали. За последнее время, может, что-то изменилось, с весны не заглядывал на их сайт. Я целиком согласен, что ветвь эта — очень молодая, и потому никак не могла сформироваться в динарском «ледниковом убежище». Говорить уверенно про 2-3 тысячи лет я бы не рискнул, но что не ледниковая эпоха, и даже не мезолит — это точно. Это не значит, что на Балканах, или где-то еще в Европе, в палеолите и мезолите не могло быть любой концентрации предковой M423. Но это значит, что прародитель всех миллионов нынешних «динарцев» тогда в каждом поколении был представлен строго одним индивидом, притом мутациям L147.2, L147.4, L343, а может даже и L621/S392 еще предстояло появиться у его весьма отдаленных потомков. Не будет гадать, где именно вилась эта ниточка на протяжении тысячелетий. Важно, что однажды она начала разрастаться. В большинстве случаев такие вещи связаны с неолитической революцией. Именно тогда сформировались зародыши современных огромных кластеров. В эпохи энеолита – бронзы они сильно перемешались, и все последующие случаи быстрого разрастания популяций приводили к росту не одной линии, а своеобразного ассорти из нескольких коррелирующих. Именно так случилось у славян: доминирует у них вовсе не одна R1a1, как по старинке думают многие, а целый «ассорти» из нескольких параллельных ветвей: M458 (на уровне от 5 до 10 %), ее дочерняя L260/S222 (от 2-3 до 10 %, а у поляков – более 13), примерно пять ветвей S466/Z280 (от 1 до 10 % каждая). В сумме это и дает знаменитые 50 и более % R1a1. Сопоставимая с ними по возрасту I2a3а-L147.2 на этом фоне – безусловный лидер, самая массовая славянская гаплогруппа, представленная в большинстве популяций на уровне порядка 10-20%, а у сербов и хорватов — выше 30%. Нас обоих интересует, когда и каким образом она влилась в славянский генофонд – ведь у предков славян в бронзовом веке, похоже, ее не было совсем (иначе она была бы не только у славян, но и у их ближайших родственников балтов). В Вашем статистическом анализе сделана попытка прояснить этот вопрос. Примем пока, что методологически там все верно и результаты заслуживают полного доверия. Суть их вы выразили картинкой со стрелками. Непонятно только одно: почему Вы называете прародину карпато-ПОЛЕССКОЙ, если у Вас четко видны стрелки, ведущие с Карпат в сторону Полесья. Если же убрать слово «полесская», то не так уж велика становится разница с моей карпато-балканской (особенно учитывая зигзаг в Словению). Вот динарским называть этот кластер действительно не стоит. В дальнейшем, чтобы не было путаницы с географическим понятием «карпатский», буду пользоваться термином «карпийский». Звучит, по-моему, не хуже, чем «арийский» smile На этом, к сожалению, трогательное единодушие кончается. Справедливо обвиняя оппонентов в недооценке разнообразия гаплотипов, Вы недооцениваете их апелляцию к относительной частоте гаплогрупп. Поясню это буквально на пальцах. Представим себе две чаши, в которых перемешаны в разной пропорции черные и белые шарики. Будем с закрытыми глазами черпать по жмене шариков из каждой чаши и высыпать в третью (это у меня такая модель смешения популяций). Суть в том, что, если шариков достаточно много и они действительно хорошо перемешаны, то Вы никогда не получите в третьей чаше концентрацию шариков одного цвета выше, чем в одной из исходных – все значения будут только промежуточными. Думаю, Вы поняли аналогию: исходная праславянская популяция с концентрацией карпийцев ниже 30 % может дать частоты, характерные для сербов и хорватов, только смешиваясь с субстратом, у которого эта концентрация гораздо выше. Это правило может нарушаться, если шарики плохо перемешаны. На одном краю чаши лежат почти одни черные, и если зачерпнуть оттуда, в третьей чаше черных окажется больше, чем в среднем в каждой из исходных. Применительно к популяциям это означает, что одна из исходных была гетерогенной. Говоря словами незабвенного Владимира Вольфовича, в числе русских там числятся и сыновья «журналистов». Притом их не просто много, а они компактно сконцентрированы в зоне будущего контакта с другой популяцией. Возможна ли такая ситуация в пражской культуре? Учитывая степень ее археологической однородности – маловероятно, но чем черт не шутит. В предыдущем посте я перечислил возможных кандидатов на роль «сыновей журналистов». Методом исключения к карпийскому кластеру могли принадлежать только потомки даков или чернолесского субстрата лесостепных скифов. Мы приходим к тому же, что Вам не понравилось в моей статье – у фракийцев концентрация карпийского кластера была порядка 40 %, и к славянам он попал именно от них. Хорваты в этом случае – почти сплошь ославяненные «журналисты», т.е. в данном случае — фракийцы. Альтернативное объяснение – высокая концентрация карпийского кластера уже сложилась в Сербии и Хорватии к приходу славян, и они ее лишь понизили, но никак не повысили. С каким этносом и с какой культурой можно в таком случае связывать родину этого кластера – можно будет порассуждать отдельно. Но к этногенезу славян это прямого отношения не имеет. Ясно, что от этого же этноса она к славянам и попала – возможно, не до, а после их дунайского расселения. Эту версию я тоже упоминал в своей статье. Если есть возражения – пишите, обсудим. Если хотите – могу пройтись и по методологии Вашего статанализа, там тоже не все просто. ********* А что касается разрастания, то Вы применяете тот тип моделирования (по методу Монте-Карло), с которого я начинал еще в 80-е. Поэтому я хорошо представляю, насколько он чувствителен к исходным посылкам. К тому же в стохастическом процессе бессмысленно говорить о темпах экспансии конкретной субклады — существует лишь распределение вероятностей от полного вымирания до предельной экспансии (вариант хромосомного Адама). Место реальной клады в этом распределении вычислить нельзя в принципе. Можно лишь показать, что данный темп экспансии при принятых допущениях возможен или исключен, только и всего.»
Ответ оппоненту и критика версии происхождения гаплогруппы I2 в изложении академической популяционной генетики:
«Вам показалось, что я — сторонник происхождения динарского кластера из «ледникового убежища». Но перечитайте внимательнее мой текст — я этого вовсе не утверждаю, пишу только о неолитических культурах на местном мезолитическом субстрате. Почему мезолитическом? Да потому, что неолит анатолийского происхождения не мог содержать I2a. Уже установлено, что его представители имели G2a и E1b-V13, продолжают дискутироваться J2 и R1b, но никак не I2a.”
По этому пункту вряд ли можно что-то возразить. Однако в целях научной педантичности необходимо сделать пару поправок. Во-первых, вопрос о том, могла ли быть I2a привнесена в Европу в ходе неолитической демагрофической революции никогда и ни кем всерьез не ставился. Статья о I2 в русской Википедии была написана любителями, и в плане содержащейся в ней информации значительно устарела. Поэтому Вы правильно сделали, что даже не стали обсуждать содержащиеся в этой статьей ложные и трудноверифицируемые выводы о том, что “носители культуры Балканского неолита (в том числе и Трипольской культуры). Последний тезис подтверждается тем фактом, что как балканский неолит является развитием ближневосточного, так и балканская гаплогруппа I родственна ближневосточной гаплогруппе J, хотя данные об их расхождении разнятся.” Тут следует сделать небольшой экскурс в прошлое. В самом начале систематического изучения вариативности человеческой Y-хромосомы в контексте масштабных популяционных передвижениях, в работах таких маститых ученых с мировым именем, как Андерхилл (2007) и Роотси(2004) была выдвинут простая, но по тем временам крайне убедительная гипотеза “трех рефугиумов”. Это теперь не вызывает никакого сомнения, что эта нашумевшая гипотеза “трех гаплогруппных рефугиумов” была частично сознательной уловкой с целью адаптации нового знания под уже устоявшиеся взгляды последователей отца-основателя популяционной генетики Л.Кавалли-Сфорца. А тогда эта новая теория была воспринята как догма, и как не прискорбно многие из популяционных генетиков продолжают ей следовать. Согласно этой теории, предки современных носителей гаплогруппы I пережили неблагоприятный период последнего ледникового периода в динарско-балканском рефугиуме. А аргументировали свое заключение отцы игрек-хромосомоведения крайне простым силлогизмом – присутствие I на Балканах во времена столь глубокой древности якобы маркировано уже тем фактом, что I и сейчас очень много. Разумеется, о дальнейшей детализации гаплогрупп и разбиении на субклады на момент написания статьи не могло быть и речи (красноречивым фактом служит хотя бы то, что гаплогруппа I2a не детализировалась ниже уровня снипа P37.2). C появилась целая группа деклассированных антропологов-подпевал, которые стали петь дифирамбы гению попгенетиков – вот мол, какой убедительный аргумент в пользу антропологической приемственности населения Балкан дали нам в руки генетики. Авторитет вышеупомянутого коллектива авторов был столь велик, что это удивительное в своей примитивности умозаключение на протяжении последущих 8-10 лет перекочевывал из одной статьи в другую, пока окончательно не оформился в виде неоспоримого догмата. Казалось бы — любое научное знание принципиально не является окончательным, а есть лишь промежуточная интерпретация истины, подразумевающая последующую замену на лучшую интерпретацию. Однако прочитав последную статью хорватских ученных о гаплогруппной вариативности населения Хорватии, я – к своему глубокому прискорбию – еще раз убедился, что мои наихудшие опасения насчет феноменальной косности научного мышления оправдались. В самых дурных традициях квазиакадемического стиля, идеологически анагажированные хорватские генетики продолжают разглагольствовать о “палеолитической балканской гаплогруппе I”. А из научно-популярной заметке (по своей сути, кратком реферате статьи) в сплитской газете благодарный хорватский обыватель узнает о том, что хорваты – это древнейший автохтонный народ Европы. Так-то!
Вячеслав Леонидович, Будем считать предыдущее сообщение пролегоменами к критике научных представлений о субкладе I2a, и поэтому перейдем к фактологической составляющей Ваших утверждений:
Итак, мы будем обсуждать происхождение так называемого динарского кластера гаплогруппы I2a.»
Во избежание возможных смысловых экивоков, сразу же предлагаю отказаться от дальнейшей «динаризации» субклады, которую мы будем обсуждать. «Динарский кластер» — это собрикет, придуманный Кеном Нордведтом для обозначения той ветви I2, которая в дальнейшем получит официальное ISOGG название I2a1b3 (L621/S392, что идентично в старой версии I2a2a). Динарский кластер целиком входит в состав этого субклада, являя собой монофилетический таксон. В профессиональной литературе этим термином никто не пользовался и никогда не будет пользоваться. Это хорошо, поскольку у непосвященного в таинства молекулярной антропологии этот собрикет может вызвать ложные аллюзии с «динарским антропологическим типом».
«Раньше его выделяли по мутации M423, но сегодня уже известно, что эта ветвь делится на дочерние, из которых нас интересует L621/S392 и ее подветвь L147.2.»
В этом предложении сразу две фактологические ошибки. Мутация M423 маркирует не I2a1b3, а ее родительскую группу I2a1b. Строго говоря, последняя делится на четыре ветви: a) гипотетический корень I2a1b* b) упомянутая в статье 2002 года I2a1b1 M359.2/P41.2 (хотя до сих пор НИКТО не видел ни одного реального гаплотипа этой группы) c) I2a1b2 L161.1/S185 d) I2a1b3 L621/S392 Две последние ветви I2a1b2 и I2a1b3 имеют строго определенный эксклюзивный ареал распространения, практически не пересекающийся друг с другом. I2a1b2 представлена, в основной своей массе, на Британских островах (отсюда и название «I2a-Isles», данное все тем же «крестным отцом» Нордведтом. Я не буду пока останавливаться на этой ветви и перейду к непосредственно интересующей нас I2a1b3 L621/S392. Здесь следует указать на Вашу вторую ошибку -| Как показало WTY-тестирование, мутация L621/S392 определяет СВОДНУЮ группу, состоящего как из ДИНАРСКОГО кластера, так и кластера DISLES (не знаю как перевести это на русский) — промежуточного между ДИНАРСКИМ и ОСТРОВНЫМ кластерами.
Ее более поздним ответвлением, видимо, являются >мутации L147.4 и L343.»
Это умозрительно. Во-первых, снип L147 [+] встречается в разных группах. Об этом я уже раньше упоминал, причем в IJ он встречается в разных субгаплогруппах аж 3 раз. Такие вотильные снипы не могут быть основанием для надежной филогении. Ergo, мы не можем говорить о том что этот снип разбивает ветвь на надежные в плане исторической интерполяции ветви. Почему существуют такие снипы, никто не смог внятно объяснить ни с позиций генетики, ни молекулярной биологии. То же самое касается L343, которые был выявлен также и у некоторых членов гаплогруппы I1.
Произошли они до формирования динарского >кластера или после — пока неясно, поскольку >определены они у считанного числа индивидов. Но >L147.2 выявлена, помимо славян, у как минимум >одного англичанина (его Kit Number в >соответствующем FTDNA-проекте — 14703), двух >немев (162426 и 164573), одного итальянца (93943) >и одного еврея из Литвы (211949), а L147.4 и L343 — >только у славян: двух поляков (209633 и 76814) и >боснийца (154978). При этом никого с L147.2+ на >L147.4 и L343 не тестировали.
Cм.выше.
Я целиком согласен, что ветвь эта — очень молодая, >и потому никак не могла сформироваться в >динарском «ледниковом убежище».
Говорить уверенно про 2-3 тысячи лет я бы не рискнул, но что не ледниковая эпоха, и даже не мезолит — это точно. Это не значит, что на Балканах, или где-то еще в Европе, в палеолите и мезолите не могло быть любой концентрации предковой M423. Но это значит, что прародитель всех миллионов нынешних «динарцев» тогда в каждом поколении был представлен строго одним индивидом, притом мутациям L147.2, L147.4, L343, а может даже и L621/S392 еще предстояло появиться у его весьма отдаленных потомков. Не будет гадать, где именно вилась эта ниточка на протяжении тысячелетий. Важно, что однажды она начала разрастаться. В большинстве случаев такие вещи связаны с неолитической революцией. Именно тогда сформировались зародыши современных огромных кластеров. В эпохи энеолита – бронзы они сильно перемешались, и все последующие случаи быстрого разрастания популяций приводили к росту не одной линии, а своеобразного ассорти из нескольких коррелирующих. » Не совсем так. Вы пишите, что в большінстве cлучаев экспансия гаплогрупп связана с неолитической революцией. Это не совсем верно. Поскольку размер эффективной популяции для Y-хромосомы в тысячи раз меньше аналогичной для аутосомных хромосом. Это означает что рост численности Y-хромосомной гаплогруппы или кластера может происходит относительно быстро и так же стремительно снижаться. Особенно если речь идет об относительно закрытом сообществе. Этот вопрос был неплохо изучен в известной статье «Y-хромосома как сигнатура гегемонии».
Уважаемый Вячеслав Леонидович! Еще раз прокомментирую Ваш комментарий:
«Итак, мы будем обсуждать происхождение так называемого динарского кластера гаплогруппы I2a. Раньше его выделяли по мутации M423, но сегодня уже известно, что эта ветвь делится на дочерние, из которых нас интересует L621/S392 и ее подветвь L147.2»
Про M423 я уже писал ранее в предыдущих комментариях. Что касается L621, то я не вижу особой филогентической полезности мутации для I2a1b3. Она (мутация) не разбивает кладу на субклады, соответствующие выявленным ранее кластерам Din-N или Din-S этой клады.Польза этой мутации может быть только в том, что он может заменить нынешную мутацию L69.2, которая была найдена и в других группах (о чем и свидетельствует цифра два после точки). Примерно год назад я обсуждал результаты теста WTY Y-хромосомы одного из представителей I2a1b3 — пана Станислава Плевако.Не вдаваясь в технические сложности, тест WTY можно описать как прочитку значительной части игрек-хромосомы. Когда я поинтересовался результатами, он сказал что сравнение результата Добсона (из «южно-динарского» кластера) и результата Плевако ( из «северо -динарского» кластера) не выявило ни одной разницы на больше 900 локусов и 215.000 нуклеотидов проведенного сиквенса игрек-хромосомы. Отрицательный результат по этому снипу у другого участника WTY Roy Hale из братской клады I2a2b Isles (L161) сузил снип L621 до 3 кластеров: южно-динарского, северно-динарского и кластера Disles («динарцы на Британских островах»). Поэтому можно считать L621 эквивалентом L69.2 — мутации, которая не отвечает критериям включения кладообразующих мутаций в официальное филогенетическое дерево ISOGG. Чуть позже результаты еще двух I2a1b3 — Пейовича из Черногории и Любинецкого из Польши (76814 Poland – Lubiniecki — I2a2 (M423+, L161-)) подтвердили верность предыдущего умозаключения. Далее Вы пишите:
Сопоставимая с ними по возрасту I2a3а-L147.2 на этом фоне – безусловный лидер, самая массовая славянская гаплогруппа, представленная в большинстве популяций на уровне порядка 10-20%, а у сербов и хорватов — выше 30%. Нас обоих интересует, когда и каким образом она влилась в славянский генофонд – ведь у предков славян в бронзовом веке, похоже, ее не было совсем (иначе она была бы не только у славян, но и у их ближайших родственников балтов)<…>На этом, к сожалению, трогательное единодушие кончается. Справедливо обвиняя оппонентов в недооценке разнообразия гаплотипов, Вы недооцениваете их апелляцию к относительной частоте гаплогрупп.»
Вы повторяете одну из главных ошибок новичков в молекулярной генеалогии, а именно основываете свои выводы на данных о частотах распространения гаплогрупп. Все делали эту ошибку, и я в том числе. Однако, как показал еще Животовский в своей известной работе: «географическое происхождение гаплогрупп Y-хромосомы можно установить по следующему эмпирическому критерию: в том месте или популяции, где возникла данная гаплогруппа, ее частота и STR-дисперсия (или возраст STR-изменчивости) максимальны по сравнению с другими популяциями (Sengupta et al., 2006). При их несовпадении (когда максимум частоты гаплогруппы приходится на один географический регион, а максимум дисперсии – на другой), место возникновения гаплогруппы становится неопределенным, но при необходимости сделать предварительное заключение предпочтение следует отдавать дисперсии как статистически и эволюционно более устойчивому показателю.» Случай с I2a1b3 -это как раз тот самый случай, когда один из пиков-максимумов приходится на Боснию-Герцеговину, а максимум дисперсии на Карпатский регион (см.мои расчеты). Следуя правилу правой руки в изложении Животовского, следует предпочесть последний вариант, так как частоты гаплогрупп больше подвергнуты флуктуациям. Далее, вторая ошибка новичков, которая встречается в Ваших тезисах, это игнорирование масштабов (Masstabe) регионов, сравниваемых по частотам гаплогрупп. Хорошо, давайте возьмем нижний порог частот по допустим, что в среднем в Польше 56% R1a1 и 10% I2a1b (хотя в зависимости от региона Польши и характера выборки статистика сильно плавает). По состоянию на 2000 год в Польше проживало 38 559 110, пусть из них половина мужчин 19279555, то 10% I2a1b это 2 млн.человек. В Хорватии население 4.496.000 или 2.24 млн мужчин, из них пусть 32% (716 800 человек будет I2a1b1). Аналогично, у сербов в среднем должно быть около 1 млн человек субклады I2a1b1. Наконец, в БиГ с пресловутым пиком I2a1b1 по разным оценкам до 60% должно проживать около 1.5 млн представителей I2a2. Вы наверное удивитесь, но я видел десятки людей приходящих на зарубежные форумы с такими же представлениями о I2a1b1. Даже ссылки давались на те же самые карты, соотношение R1a1/I2a (который один из «мудрецов» на DNAforums обозначил как коэффициент славянскости). Правда, практически все они были убеждены в конце концов в моей правоте. Какими способами можно определить гипотетический расклад гаплогруппа у исторических и доисторических народов? Существует три апробированных способа. I.Первый способ — самый надежный, так как базируется на материальных доказательствах (то есть древней ДНК, извлеченнной из археологических специменов). Какое новое знание — о распределении гаплогрупп на Балканах в течении последних нескольких тысяч лет — может дать этот метод? Исходя из данных о «ископаемых» Y-гаплогруппах в неолите (включая генотипирование знаменитой мумии Отци), можно уверено предположить, что они были представлены гаплогруппами G2a, E1b, J2b, J2a, R1b1b2 — то есть теми гаплогруппами, которые связываются с цивилизациями «старой Европы» (с) Гимбутас и более широко с неолитической демографической революцией и миграциями времен бронзового века в Европе.Я же полагаю, что «иллирийцы» тех времен были prima facie представителями субклад J2a1, J2b,E1b1b1a (особенно E1b1b1a2 (E-V13)), субклад G2a (особенно G2a3a), субкладов R1b1b2 (особенно R1b1b2a1a2d) и так далее. Ядро гаплогрупп балканских гаплогрупп времен бронзы по мере убывания могло выглядеть следущим образом:E1b1b1a2, J2b2, J2b1, G2a3, R1b1b2a1a,J2a4, J2b1. Последние работы по анализу древнего ДНК извлеченных из останков в неолитических захоронениях только подтвердили эту гипотезу — там в основном были E1b1b и G2a. То, что сейчас у югославов эти группы в меньшинстве явно объясняется мощным «бутылочным горлышком» времен аварского каганата. II. Второй способ — состоит в интерполяции состава и распространения гаплогрупп в изолированной «потомковой популяции» на распространение гаплогрупп в предковой популяции. Этот способ был использован в недавней статье о гаплогруппах арберешей — потомков албанцев, мигрировавшей в средние века в Италию, где они образовали изолированную группу. Чтобы освежить Вашу память, напомню, что албанцы считаются прямыми потомками иллирийцев. На основании анализа выборки было показано, что вышеупомянутые неолитические балканские гаплогруппы у арберешей по своему распределению соответсвуют современному раскладу этих гаплогрупп на Балканах. В то время как %I2a у арберешей гораздо ниже как среднего % I2a у албанских популяций, так и средней частоты распространения I2a на Балканах. О чем это говорит? О том, что I2а — (относительно) недавние пришельцы на Балканах. Далее, оценка возраста «югославских кластеров» показывает, что они все не «старше» 1400-1500 лет до настоящего времени, что означает бурную экспансию субклада в 5-6 веках н.э. III Третий cпособ -наиболее уязвимый, т.к. строится на филогенетической и статистической обработке массива гаплотипов, оценки возраста БОПа, оценки молекулярного разнообразия, оценки структуры и качества филогенетического дерева и т.д. Я построил десятки тысяч деревей субклада I2a2, а также оценил молекулярное разнообразие субклада I2a2, который оказался близким к % разнообразия из статьи Перчич. Структура дерева, в котором «северные» I2a2-Din находятся ближе к корню, чем «югославы», — а также более высокое молекулярное разнообразие I2a2-Din на севере Восточной Европы достаточно точно свидетельствуют о том, что прародина I2a2 находилась именно в это месте.
Ответ Вячеслава Носевича:
Я не буду в очередной раз комментировать Ваши опровержения моих якобы ошибок, которые происходят от недопонимания того, что я пытался сказать. Это не затрагивает сути обсуждаемой темы. А в ней я вижу два важных аспекта, свою позицию по которым хочу прояснить.Первый аспект связан с определением возраста клады по разнообразию гаплотипов. Почему он оказался столь ненадежным, что Диенекес, например, вообще в нем разочаровался? Дело не в математике как таковой, и не в скорости мутаций. Дело в особенностях ветвящегося случайного процесса, который это разнообразие порождает. Его простота обманчива. На начальной стадии роста клады он легко моделируется по методу Монте-Карло. Клада растет практически независимо от динамики популяции в целом, а если популяция невелика и относительно стабильна (в каменном веке они почти все были такими), то рано или поздно численность «удачливой» клады приближается к эффективной численности популяции. Это – предельный случай генетического дрейфа, когда клада заполняет чуть ли не всю популяцию (реальные примеры описаны у малых народов Сибири, в горных долинах Дагестана, частично этому соответствует и понравившаяся Вам статья про О’Нейлов). Гаплотипы при этом образуют четкий «стар-кластер», в котором модальным является исходных гаплотип разросшейся клады. Соответственно и ее гаплогруппа абсолютно доминирует. Зная скорость мутаций в локусах и измерив частоту гаплотипов, мы можем точно определить возраст клады.А дальше – самое важное. Рост численности разросшейся клады и популяции в целом перестает быть независимым – это очевидно. Что произойдет, если популяция вдруг начнет расти, и даже распадется на несколько дочерних, относительно изолированных? Разросшейся кладе расти уже некуда, начинается рост дочерних. А поскольку наиболее велика частота гаплотипов, близких к исходному (это ощущается тем сильнее, чем меньшее число локусов мы учитываем), то большинство этих дочерних клад имеют именно такие гаплотипы основателей. Все они порождают новые стар-кластеры, которые почти целиком перекрываются и внешне сливаются в один. Число маргинальных гаплотипов становится аномально низким, а близких к исходному – аномально высоким по сравнению с предсказанием модели, предполагающий стабильную численность популяции или равномерный рост.Все. Теперь сколько бы мы ни измеряли частоту гаплотипов и в какие статпакеты ее ни засовывали, получаемый возраст будет фиктивным (заниженным). А если рост популяций будет пульсирующим (по циклу разрастание – остановка=дрейф – разрастание), то в большинстве из них будет тысячелетиями воспроизводиться исходный стар-кластер, и лишь в некоторых его основой могут стать маргинальные гаплотипы, сильно удалившиеся от исходного.Хотите реальные подтверждения? Самое яркое – гаплогруппа R1b1. Кластеры U106 и P312, разошедшиеся порядка 10 тыс. лет назад, до сих пор практически неразличимы по частоте гаплотипов, а модальные крайне близки что по 67 локусам, что по 102. Столь же близок модальный у предковой L11*, от которой они отпочковались еще раньше. А вот одна из дочерних клад, ирландская М222 (по-моему, именно на нее наткнулись исследователи О’Нейлов) образовалась вокруг одного из маргинальных гаплотипов. По любому методу расчетов точка ее ответвления окажется старше, чем точка расхождения U106 и P312, хотя реально она примерно вдвое их моложе!Аналогичный случай – с индийской R1a1-Z93 и европейской R1a1-Z83. Разошлись они, по моим оценкам, примерно 9 тыс. лет назад, а столь «обожаемый» Вами Клёсов вычислил вдвое меньший возраст. А на примере с E1-V13 погорел Диенекес (как и тот же Клёсов), когда ее возраст в ископаемой ДНК тоже оказался вдвое старше расчетного.Если есть мнение на этот счет – высказывайте. А про второй аспект напишу в следующий раз.
А что тут комментировать? Описанные Вами феномены хорошо известны всем исследователям — как профессиональным попгенетикам, так и любителям. Вы еще забыли добавить сюда полный раздрай в плане определения скоростей мутаций (об этом писал еще почти 10 лет назад Животовский в своих основопологающих статьях о микросателлитной изменчивости и эволюционных скоростях мутаций). Но все это относится к вопросам определения TMRCA и опосредованно к определению дисперсии.Филогенетический анализ основан на других принципах, и по этому Ваше глубокое замечание интересно, но имеет отдаленное отношение к предмету дискуссии.
Теперь, уважаемый Вячеслав Леонидович, перейдем к анализу НАИБОЛЕЕ важного (как мне представляется) аспекта Вашей теории:
Непонятно только одно: почему Вы называете прародину карпато-ПОЛЕССКОЙ, если у Вас четко видны стрелки, ведущие с Карпат в сторону Полесья. Если же убрать слово «полесская», то не так уж велика становится разница с моей карпато-балканской (особенно учитывая зигзаг в Словению). Вот динарским называть этот кластер действительно не стоит. В дальнейшем, чтобы не было путаницы с географическим понятием «карпатский», буду пользоваться термином «карпийский» <…> Думаю, Вы поняли аналогию: исходная праславянская популяция с концентрацией карпийцев ниже 30 % может дать частоты, характерные для сербов и хорватов, только смешиваясь с субстратом, у которого эта концентрация гораздо выше. <…> Возможна ли такая ситуация в пражской культуре? Учитывая степень ее археологической однородности – маловероятно, но чем черт не шутит. В предыдущем посте я перечислил возможных кандидатов на роль «сыновей журналистов». Методом исключения к карпийскому кластеру могли принадлежать только потомки даков или чернолесского субстрата лесостепных скифов. Мы приходим к тому же, что Вам не понравилось в моей статье – у фракийцев концентрация карпийского кластера была порядка 40 %, и к славянам он попал именно от них.Хорваты в этом случае – почти сплошь ославяненные «журналисты», т.е. в данном случае — фракийцы. Альтернативное объяснение – высокая концентрация карпийского кластера уже сложилась в Сербии и Хорватии к приходу славян, и они ее лишь понизили, но никак не повысили. С каким этносом и с какой культурой можно в таком случае связывать родину этого кластера – можно будет порассуждать отдельно. Но к этногенезу славян это прямого отношения не имеет. Ясно, что от этого же этноса она к славянам и попала – возможно, не до, а после их дунайского расселения. Эту версию я тоже упоминал в своей статье.»
На мой любительский взгляд «карпийская часть» Ваших рассуждений представляет собой наиболее уязвимую часть Вашей внешне стройной концепции. Я намерено выношу за скобки нашей дискусии Ваши примеры с «журналистами» и «шариками». Я также не буду обсуждать здесь корреляции с археологическими культурами, языками, а также теорию балто-славянской общности. Во-первых, я считаю себя недостаточно компетентным в этих вопросах, а во-вторых — это займет слишком много места. Скажу лишь что проблема грамотной аппроксимации результатов генетических исследований и всего огромного массива наработок лингвистики, этнографии и археологии по вопросу этногенеза славян — это дело будущего и потребует много лет вдумчивого системного анализа. Что касается присутствия в названии слово «полесский», то оно носит арбитрарный характер, и вовсе не означат что предковая популяция I2a1b3 «существовала» изначально в Полесье. По моим скромным представлениями она вышла на популяционный уровень где-то между Карпатами и Полесьем. Отсюда и двойное название. Я лишь останавлюсь на тех моментах, которые мне представляются наиболее спорными:
- 1) тезис о (гаплогруппной) однородности протославян — мне кажется что это очень слабый тезис, так как последние исследования генофонда неолитических культур Европы показали, что они были далеко неоднородны. Элементарная логика подсказывает, что в бронзовом веке и позднее степень смешения должна была только увеличится. Поэтому возможны альтернативные точки зрения. Например, задумывались ли Вы о том, что «ассимиляция» или «слияние» общностей представителей кладов R1a и I2a1b3 могла произойти еще до времени возникновения «славянской» общности.
- интерполяция современных частот гаплогрупп — манипуляции с современными частотами в целях реконструкции гипотетических частот распространения в генофонде древних народов также вызывают закономерные вопросы. По-крайней мере, подобные эксперименты возможны только исходя из генетических данных полученных в результате анализа останков из захоронений соотвествующих культур или народностей. В противном случае расчеты совершено произвольны, и им нельзя доверять. Вот наглядный пример. В 2011 году вышла интересная статья «Linking Italy and the Balkans. A Y-chromosome perspective from the Arbereshe of Calabria.» Boattini A, Luiselli D, Sazzini M, Useli A, Tagarelli G, Pettener D. В работе исследовалась популяция арберешей в Калабрии. По мнению авторов игрек-хромосомный генофонд арберешей должен отражать состояние игрек-хромосомного предкового генофонда алабанцев 500 лет назад. В числе прочего авторы приходят к интересным выводам: » Intra-haplogroup analyses suggest that this area may have experienced important changes in the last five centuries, resulting in a marked increase in the frequency of haplogroups I2a and J2.» Предложенная в статье интерпретация популяции Arbereshe как архетипа (proxy) первоначального албанского населения приводит нас к выводу, что ветвь I2a1b3 встречался в южной части Балканского полуострова 500 лет назад гораздо реже, чем сейчас. Как Вы видите — в ходе дальнейших миграций %-ная доля нескольких групп не только не уменшился, а наоборот, увеличился.
- тезис о связи доисторических носителей I2a1b3 с фрако-дакийцами — также представляется мне совершено надуманным. Признаюсь, Вы не первый кто постулировал такой тезис — еще 2 года назад я встречал подобные рассуждения на международных форумах и обычно такие идеи озвучивали румыны и некоторые болгары. Говорили о положительной корреляции ареала I2a1b с фракийским племенами, в том числе костобоками и карпами. Были и альтернативные суждения о связи с миграциями бастарнами и пр. Слабость таких рассуждений очевидна — даже если мы будем исходить из реконструкции предковых частот, то будем вынуждены признать, что I2a1b вряд ли могла присутствовать в столь значительных количествах у фракийцев, так как у болгаров и румын как частоты распространения, так и уровень дисперсии гаплотипов I2a1b значительно ниже чем у тех же словаков и западных украинцев.
Вячеслав Носевич:
«задумывались ли Вы о том, что «ассимиляция» или «слияние» общностей представителей кладов R1a и I2a1b3 могла произойти еще до времени возникновения «славянской» общности.» Не только задумывался, но и описал 3 года назад как один из возможных вариантов проникновения I2a1b: венетский клад слился с уже трижды ассимилированными до того потомками чернолесцев (первый раз — они стали скифами, второй — милоградцами-неврами, третий — бастарнами). Собственно, с Вашего несогласия с этим пунктом и началась наша дискуссия. И теперь вы предъявляете его мне как опровержение меня… Но меня пока еще не убедили, что этот компонент складывающегося славянства достаточен для объяснения наличия I2a1 у румын и хорватов. Вот если разберемся с дисперсией в сербохорватской и белорусо-украино-польской частях кластера, может, куда-то и продвинемся. А пока у меня ощущения, что дискуссия начала ходить кругами.
Уважаемый Вячеслав Леонидович! Вы пишете:
«Хорваты в этом случае – почти сплошь ославяненные «журналисты», т.е. в данном случае — фракийцы.» Это весьма сильное заявление и нуждается в подробном анализе. Я воспользуюсь свомими заметками и заметками некоторых активистов форума Молген, в частности уважаемого Вячеслава Малиновского. В известной статье Перичич опубликованны данные только по I-37 в целом , причем в качестве исследованных регионов у Перичич были указаны материковая Хорватия и четыре острова, а не Долмация в целом. Если же вы говорите, что все хорваты к примеру это «почти сплошь ославяненные «журналисты», т.е. в данном случае — фракийцы, то по идее у них должны быть примерно тот же расклад по гаплогруппам что и например у болгар. Например, y герцеговинских хорват 71,1% I2a2, в отличии от местных сербов -31% и босняков 43,5%. И пресловутой R1b там у хорватов 2%, а у сербов 6%, а вот Е у сербов как и везде за 20%, а у хорватов 9%(больше чем в других хорватских регионах, но все-таки в 2 раза меньше чем у сербов). У болгар: 16% E1b1b 1% G2a 3% I1 20% I2a 1% I2b 20% J2 1% Q 18% R1a 18% R1b 1% T Кроме того, болгары в отличие большинство других балканцев (в первую очередь хорватов) преимуществено I2a1b3-«cеверные динарки». В этом смысле они гораздо ближе полякам и украинцам.
Далее, Вы пишете, что:
«Альтернативное объяснение – высокая концентрация карпийского кластера уже сложилась в Сербии и Хорватии к приходу славян, и они ее лишь понизили, но никак не повысили. С каким этносом и с какой культурой можно в таком случае связывать родину этого кластера – можно будет порассуждать отдельно. Но к этногенезу славян это прямого отношения не имеет. Ясно, что от этого же этноса она к славянам и попала – возможно, не до, а после их дунайского расселения. Эту версию я тоже упоминал в своей статье.»
Это интересное, но весьма спорное утверждение, так как оно предполагает некое автохтонное присутствие I2a1b3 на территории Сербии и Хорватии к приходу славян. Тут сразу возникает ряд несостыковок. Если под автохтоностью I2a2 на Балканах подразумевается размещение «прародины» этой гаплогруппы на Балканах, то я категорически несогласен. В противном случае, я вполне согласен рассматривать югославских I2a2 как автохтонов Балкан, с той лишь оговоркой, что эти «автохтоны» появились на Балканах не ранее 6-7 веков н.э., вскоре после того как авары «зачистили» местное автохтонное население. Лично для меня совершенно очевидно, что по Y-гаплогруппному составу население Далмации,Иллирика времен римского господства коренным образом отличается от населения этих же земель после славянского завоевания. Между прочим, вот Вам интересный факт для размышления. Историкам хорошо известен период римской истории, который носит название период иллирийских императоров. Формально он начинается от смерти Галлиена (268 г. н. э.) до начала правления Диоклетиана и предшествует периоду домината. На самом деле же сам Диоклетиан, а также пришедшая к власти династия Флавиев, так же как и ряд позднейших императоров были выходцами из Иллирика, Паннонии, Далмации, Сирмии, Мезии( это нын.Хорватия,Босния,Сербия,Болгария). Очевидно, что кроме императоров и солдат, эти провинции поставляли в Рим и гражданскую администрацию, и самое главное — огромную массу обслуживающего персонала). Известно также, что иллирийские императоры (правившие Римом в 3-4 веках), подобно современным албанцам, черногорцам и т.д, практиковали клановую систему. Известно множество случаев, когда после головокружительной карьеры и прихода к власти после военных переворотов, эти парни из балканских деревушок перевозили в Италию своих родственников целыми кланами и деревнями. А теперь внимание, вопрос знатокам — ежели мы допускаем, что локальный пик частотного распределения гаплогруппы I2a2 на Балканах существовал в неизменном состоянии испокон веков (хотя бы со времен LGM), то логично было бы допустить наличие подобных локальных пиков частотности I2a2 и в самой Италии. Почему же мы его не видим? (про сардинскую группу I2a1 просьба в ответе не упоминать, так как это отдельная песня). С одной стороны имеем Боснию-Герцеговину с максимальным пиком % гаплогруппы I2a1b3. C другой стороны, по историческим источникам выходит, что эти страны запустели под (например под, аварским правлением и были заселены вновь прибывшими с Карпат славянами. С другой стороны — по мнению попгенетиков, I2a1b3 все время проживали на этой территории со времен последнего ледникового максимума. По моему мнению, последние явно ошибаются.
Комментарий Вячеслава Малиновского:
Еще один момент. По сути мы обсуждаем возможность того, что славяне переселившись на Подунавье (Балканы здесь не так важны, поскольку исходной точкой расселения славян по Восточной Европе указано Подунавье), в течении 100-200 лет обновили свой генофонд где-то наполовину. Мы не берем пиковые показатели I2a2 для Боснии, а рассматриваем таковые для Болгарии и Румынии, Венгрию тоже не вижу ставить в этот ряд — по генофонду это практически родные братья словаков и чехов. Итак даже отбросив R1a1 в Румынии и Болгарии (считая что якобы все клады этой гаплогруппы были принесены славянами, хотя это конечно же не так), получим I2a2 в этих странах на уровне 30-40%, остальных гаплогрупп 60-70%. Теперь риторический вопрос — ассимилировав в рекордные сроки придунайское население, славяне ведь не могли принимать в свою среду мужчин исключительно с гаплогруппой I2a2? Следовательно кроме нее славяне должны были «взять» и другие местные гаплогруппы, в ДВА раза больше чем I2a2. Получаем, что у славян Восточной Европы кроме 15-20% I2a2 должны наличествовать еще 30-40% иных «балканских» гаплогрупп. Где они?
Вячеслав Носевич:
Вячеславу Малиновскому: Хороший вопрос. Попробую перефразировать его: славяне в ареале пражской культуры имеют соотношение гаплогрупп I2a1 и R1a1 примерно в соотношении 1 к 2. Если они принесли I2a в Хорватию и Сербию, то вместе с их 33-36% должно было появиться и вдвое больше (т.е. 66-72%) R1a1. Где они? А теперь отвечу по существу. Одна из популяций, поглощенных славянами в Балкано-Карпатском регионе или ранее, еще до распространения пражской культуры (как Вы помните, это один из рассматриваемых мной вариантов), причем поглощенная полностью, без остатка, имела очень высокую концентрацию I2a1, а остальное составляли понемножку G, E, J и, возможно, R1a и R1b. Концентрация этих остальных гаплогрупп была, скажем так, типичной для региона, и на уровень у славян этот приток не повлиял. А вот аномально высокое значение I2a1 соответственно повысило его у славян.
Комментарий Вячеслава Малиновского:
Балкано-Карпатский регион это название, территориально охватывающее чуть не треть Европы. Предположение с какой-то стопроцентно поглощенной популяцией выглядит более чем натянуто, потому что мы должны придти к выводу, что эта самая популяция населяла не все Подунавье-Балканы, а всего лишь какой-то достаточно ограниченный ареал, из которого собственно как из единого центра и начались разнонаправленные славянские миграции. ТО есть и славяне здесь предстают каким-то одним централизованным, достаточно немногочисленным племенем, живущим уже не родо-племенным обществом, так как I2a2 оказалась сразу во всей славянской популяции. Где этот регион из которого начались славянские миграции? Насколько помню нулевая фаза Пражской культуры это как раз Полесье, так какие же Балканы.Если они принесли I2a в Хорватию и Сербию, то вместе с их 33-36% должно было появиться и вдвое больше (т.е. 66-72%) R1a1.» Почему? Я ведь не говорю о славянстве как генетически едином целом. Это родо-племенное общество, в каждой из групп которого свой гаплогруппный состав, что мы видим у современных народов, еще сохранивших память о своей родовой структуре. Что до R1a1 то считаю совершенно неправильно всех их относить к славянам. Например если вы наложите карту М-458 на карту пшеворской культуры, то увидите что основной массив гаплотипов находится как раз в ее ареале. И если посмотреть гаплогруппный состав чехов например, то утверждения что славяне продвинулись на земли практически полностью оставленные германцами, выглядят мягко говоря преувеличенными. То есть процент славянских мигрантов и их племенной состав в каждом регионе был различен и в одном случае местное население имеет «балканские» гаплогруппы, в другом «кельто-германские». Разный процент той же I2a2 (от 10 до 30%) говорит всего лишь об участии разных племенных группировок славян в миграциях. Что до Хорватии, то Вадим ведь указывал, что запредельные пики это показатели герцеговинских хорватов, на с-в Хорватии ситуация совсем иная. Какова величина популяции герцеговинских хорватов? Скорее всего это лишь эффект основателя.
Вячеслав Носевич:
Насчет пиков как следствие эффекта основателя — соглашусь. Но общее плато, над которым поднимаются эти пики, на Балканах выше, чем на славянской прародине. Не могли славяне принести большую концентрацию «черных шариков», чем имели изначально. В палеолите такое возможно, а в средневековье, когда перемещаются массы в сотни тысяч и миллионы людей — уже нет.Я ведь не говорю о славянстве как генетически едином целом.» Если Вы читали все комменты, то именно этот вариант я рассматривал как случай с плохо перемешанными шариками. Но род, несущий высокую частоту I2a, не мог быть исконно славянским — это я и имел в виду под «журналистами» (правильно было бы — «юристами», подзабыл я высказывание Вольфовича…)
Комментарий Вячеслава Малиновского:
Но общее плато, над которым поднимаются эти пики, на Балканах выше, чем на славянской прародине.» А что вы надеялись увидеть на территории к примеру Украины, практически по новой заселенной в 17 веке? Вот и видим пики в местах убежищ — в полесье, волынском и белорусском.Но род, несущий высокую частоту I2a, не мог быть исконно славянским» Конечно не мог, праславянский язык это какая-то из клад R1a1, но с ними точно еще не скоро получится разобраться. I2a к тому времени скорее всего уже не раз сменили язык, но я веду речь о том, что к моменту начала миграций они уже входили в славянскую общность и на данный момент это самый удобный славянский маркер — нет ни малейшего локального всплеска, который бы не объяснялся «славянским следом»-«Не могли славяне принести большую концентрацию «черных шариков», чем имели изначально. В палеолите такое возможно, а в средневековье, когда перемещаются массы в сотни тысяч и миллионы людей — уже нет.» Почему? О каких сотнях тысяч мы говорим? Пару-тройка сотен тысяч мужчин это популяция герцеговинских хорватов в 21 веке, если говорим о 6-7 веках, то там речь шла о сотнях, если не десятках. Это мог быть всего лишь один из славянских родов с высокой долей I2a.
Ув. Вячеслав Носевич, Вы наверняка спросите, а каково мое личное мнение на счет происхождения I2a1b3. Чтобы ответить на этот вопрос, необходимо начать издалека. По моему скромному разумению маршрут путешествия I2a1 начинается где-то в Альпах (примерно 12 000 -10 000 лет назад).Скорее всего, ко времени отступления ледников, «популяции» I2a1*, I2a1a, I2a1b и I2a1c уже разделились. Одни пошли на юг, положив начало иберийско-сардинским I2a1a, другие -I2a1b — на север, где в свою очередь разделились на предковую линию I2a1b3 и предковую линию I2a1b2 (последние, видимо, оказались изолированными вместе с частью I2a2, I2a1a на Британских островах после затопления Доггерленда). Предковая линия I2a1b3 же двигались постепено на северо-восток за отступающим ледником. Спустя много тысяч лет значительная часть субклада I2a1b3 «вернулась на юг» уже в составе славянских племен, поселившихся на Балканах между 6-8 в.н.э. Таким образом, значительное количество I2a1b3 совершило движение по часовой стрелке — в конечном итоге вернувшись в места, близкие к точке первоначального исхода их предков. Примерно полтора года назад удалось немного подискутировать с администратором польского FTDNA проекта (Лоуренсом Майкой) по вопросу о происхождении гаплогруппы I2a1b (и I2a1 в целом). Майка предложил крайне интересную версию места происхождениия гаплогруппы I2a1b. По его мнению, общий предок I2a1b (как I2a1b2, так и I2a1b3) c выской степенью вероятности мог жить в районе Альпийских гор. Он также согласился с предложенным мною маршрутом перехода клана предков I2a1b3 из Альп в Карпаты. Правда, он не согласен с моей датировкой. Я считаю, что предковая популяция I2a1b3 начала свое движение из Альп в сторону Карпат сразу после окончания ЛГМ (т.е. в мезолите), а Майка считает что инициатором миграции могли быть кельты (т.е эта миграция произошла уже в бронзовом веке). Последний вариант хоть и интересный, но не объясняет времени и места разделения островного субклада I2a1b2 и динарского субклада I2a1b3, которая по молекулярной датировке не могла произойти позднее чем 10-8 тысяч лет тому назад. С целью проверки версии Майки, я решил использовать алгоритм Мескита, который позволяет производить оценку наиболее вероятного (с точки зрения парсимонии) места «основания» субклада, исходя из нынешней географической дистрибуции (географического распространения) двух смежных субкладов (в данном случае мы определяем место появления I2a1b1 на основании сопоставления географических координат таксонов-гаплотипов и топологии дерева I2a1b2, которое укоренялось аутгрупп-корнем I2a1a). Несмотря на то, что наибольшая плотность субклада I2a1a приходится на Сардинию, cледут опираясь на хорошо аргументированный и эмпирически подтвержденный аргументНордтведта признать, что самые старые по возрасту кластеры I2a1а приходятся на северный регион Пиренейского полуострова, и частично южную Францию. Поэтому в качестве координат аутгруппа-корня (гаплотипа I2a1а) были приняты координаты географического центра Пиренейских гор Построенные таким образом в Меските филогенетические деревья прямиком были спроицированны на географическую карту Северной Европы, в 2D и 3D проекциях. Если хотите, то ямогу 3 карты, построенные мной в Меските-Картографере на основании структуры филогенетического дерева из 687 гаплотипов I2a1b3, выявленной путем нахождения 20 независимых совпадений 20 парсимонически лучших (наиболее оптимальных) деревьев в TNT. Это дерево было выгружено из ТНТ в формате .tre и загружено в картографический пакет Мескита, где оно было спроецировано на карту Европы (проекция Меркатора). Нанесены номера ветвей (чем дальше от корня, тем болше номер корня -см.номера в овале) и парсимонически предпологаемые маршруты миграции представителей этой генетической ветви в Европе. На примере второго «картографического» дерева I2a1b3 показана другая интересная функция картографического пакета Mesquite. А именно, наличие встроенного алгоритма для определения максимально вероятной (в терминах парсимонической комбинации географически близких ныне живущих таксонов 🙂 предковой локации субклада I2a1b3. На реконструированной карте Мескит довольно уверенно определяет место происхождения субклада где-то к северо-востоку от Карпат (обозначено большим красным кругом с красной точкой в центре).
Впрочем, на этот счет существует альтернативная точка зрения. Ув. Вячеслав Малиновский в комментариях к карте Нордведта пишет: «Исходный регион миграции I2a2-Dinaric-M423 не совсем верно очерчен (где-то к востоку от Варшавы), но направление верное — с севера на юг. Принимая во внимание приблизительный возраст субклада I2a1b3 (ок.2500-2000 лет) и разветвление дерева на уровне 1500-1700 лет (чему соответствует демографический процесс расселения), единственней миграцией такого массового масштаба можно считать именно славянскую колонизацию Балкан. Насчет возраста I2a1b3 все гуру единодушно сходятся к 2700-2500 годам. Даже сверхсекптичный Диенек Понтик считает, что I2a1b3 слишком молодая ветвь, чтобы быть кандидатом на неолитическое население Балкан. Ну как же не видите? Как верно заметил Вадим, точку возникновения L-147 по-хорошему нужно сместить несколько восточнее, в треугольник Ю.Польша-Прикарпатье-Полесье. Вот там где-то 2500 лет назад и возник I2a1b3, а далее его миграции штрихованными линиями, и по направлениям и по времени полностью совпадающие со славянскими миграциями.У I2a1b3 нет каких-то региональных ветвей, примерный возраст тех что постарше где-то 1500 лет, на них вперемешку русские, украинцы, поляки, сербы, восточные немцы. Что коррелирует со сведениями о разнонаправленных миграциях каждого из племен. К примеру хорваты частично остались на Волыни, частично пошли на Эльбу, частично переселились на Балканы. Первоначально I2 были носителями каких-то палео-европейских языков, вполне вероятно что в неолите если не все, то часть из них перешли на языки неолитических переселенцев из Малой Азии. Но например I2a1b3 не были у трипольцев коренным элементом (их ядро составляли выходцы с Ближнего Востока), а скорее потомками местного мезолитического элемента, инкорпорированного в состав этой культуры. Если говорить об исконной культуре древних I2a1b3, то это скорее всего были свидерцы, и их эпигоны.К моменту начала протославянского этногенеза субклад I2a1b3 целиком находился в ареале Лужицкой культуре — одной из культур полей погребений, относившейся к кельто-германо-италийской лингвистической общности. Отмечается что на этапе своего вычленения из общей балто-славянской лингвистической общности славянский язык испытал влияние какого-то кентумного языка, но скорее всего не собственно кельтским, ни германским этот язык не был. Я не вижу другого варианта, кроме как взаимодействие балто-славян Поморской культуры(скорее всего антов нашей классификации, позднее населения восточного ареала Пшеворской культуры)) с населением Лужицкой культуры. Далее это население частично участвовало в миграциях на восток, поучаствовав в сложении тамошних культур.»
Комментарий Вячеслава Малиновского:
Уточню — касательно свидерцев и прочих «неолитов — мезолитов» Юго-Восточной и Центральной Европы, говорить наверное можно о I2*, что подразумевает как предковые клады, так наверное и нынешнюю I2c, «размазанную» тонким слоем по всей Европе с локальным всплеском в Закавказье. Куда по моему разумению она с Балкан попасть не могла иначе, кроме как с предками армян (одними из, которые были родственны фригийцам). Что до «антов» в цитате выше, то они не имеет никакого отношения к историческим антам, это всего лишь условное и часто вводящее в заблуждение название одной из ветвей R1a1 когда-то принятое на Молгене — давно пора сменить название, но ничего «навека» никому в голову что-то не приходит.
Вячеслав Носевич:
дополнение к сказанному мной ранее: пример с R1b1 вообще-то еще более впечатляющ. Я выбрал из баз FTDNA все гаплотипы на 67 и более локусов, определенные как L150+ L51-, L23+ L51- и M269+ L150-, а заодно все с М269+ U106- P312-. Все эти ветви последовательно отпочковывались на отрезке примерно от 20 до 10 тыс. лет назад. Так вот, их сегодняшние гаплотипы по-прежнему близки до полного смешения, а модальные различаются друг от друга на считанных локусах. Например, на 17-локусном наборе Y-Filer модальные гаплотипы кластеров M269+L23- и L23+L51- не совпадают на 1 шаг по 4 локусам, тогда как Z196 и DF19 не совпадают по 5, хотя разошлись тысяч на 10 лет позже! И главное, что никаким поправочным коэффициентом этот эффект компенсировать невозможно.Почему в такой ситуации я все же уверен, что карпийский кластер – молодой (хотя, возможно, и не столь молодой, как кажется)? Ситуация с определением возраста по гаплотипам все же не совсем безнадежна. Описанный мной эффект препятствует расхождению модальных гаплотипов и снижает число маргинальных, но не запрещает маргинальным расходиться все дальше. Если бы у нас были надежные выборки, порядка сотен длинных гаплотипов в каждой ветви, тестированных по всем Y-снипам, мы увидели бы, что САМЫЕ УДАЛЕННЫЕ ОТ МОДАЛЬНОГО различаются тем сильнее, чем древнее кластер. Проблема лишь в том, что такие выборки появятся очень нескоро. Но очень грубые прикидки можно сделать даже по имеющимся данным.Результат будет надежнее, если учитывать не только генетические расстояния, но и мутабельность локусов. Ясно, что различие на 3 повтора в DYS392 значит гораздо больше, чем такое же расстояние в DYS439. Поэтому я умножаю расстояния на поправочные коэффициенты, пропорциональные известной скорости мутаций в каждом локусе. При таком подходе три самые удаленные из известных 67-локусных гаплотипов карпийского («динарского») кластера отстоят от модального на 22.9, 19.5 и 18.5 «калиброванных» шагов. В западнославянском кластере R1a1-L260 самые удаленные отстоят на 19.5, 19.4 и 18.5, а в его предковом М458+ L260- – на 36.8, 35.1 и 33.5. Рискнем предположить, что карпийский кластер близок по возрасту к R1a1-L260 (но все же чуть старше его) и существенно моложе R1a1-М458.Здесь уместно упомянуть обещанный второй аспект. Он касается репрезентативности выборок и длины использованных гаплотипов. Оценка репрезентативности – это азы матстатистики, на эту тему есть уйма учебников. Общеизвестно, что при социологических опросах применяются выборки порядка 1500 человек. Для мужской Y-хромосомы разброс значений сопоставим с разбросом общественного мнения, а эффективный размер ниже, поэтому можно примириться с национальными выборками порядка сотен, но уж никак не десятков индивидов. В этой связи посмотрите на размеры национальных выборок, использованные в Вашем анализе, и сделайте выводы…Размер тоже имеет значение (в смысле, длина гаплотипа). Клесов не раз с гордостью подчеркивал, что его метод одинаково работает на гаплотипах любой длины. Но это не достоинство, а наоборот. Адекватный метод ОБЯЗАН на длинных гаплотипах давать более точные результаты, чем на коротких (если требуется дополнительная аргументация – поясню). В этой связи использованный Вами «компот» из гаплотипов разной длины меня, мягко говоря, не впечатляет. Немножко напоминает среднюю температуру по больнице…Посмотрим, что дает предложенный мной метод при уменьшении числа локусов. Тот же карпийский кластер в варианте Y-Filer (17 локусов) дает максимальные расстояния в 7.3, 6.7 и 6.5 шагов, причем это НЕ ТЕ гаплотипы, которые показали наибольшее удаление на 67 локусах (для них результаты соответственно – 5.4, 2.7 и 4.7). Самые удаленные гаплотипы кластера R1a1-L260 теперь дают удаление 5.6, 5.1 и 5.0, причем лишь один из них – тот же, что и на 67 локусах. Два других теперь дали значения 3.5 и 0.3 (!). При этом на 17 локусах западнославянский кластер R1a1-L260 выглядит ощутимо моложе карпийского, что вряд ли верно.Мораль: даже одна и та же хромосома может дать очень разный вклад в суммарную статистику в зависимости от того, какие ее локусы учитываются. Именно поэтому так важна величина выборки, при которой эти случайные броски компенсируются законом больших чисел.Урезание именно до 17 локусов было предпринято мной, чтобы расширить выборку за счет гаплотипов из YHRD и других источников с аналогичным набором локусов. В частности, у нас есть данные из (Mirabal et al., 2010 Human Y-chromosome short tandem repeats…) с 404 черногорскими гаплотипами и 179 сербскими, из (Ljubković et al., 2008 Y-chromosomal Short Tandem Repeat Haplotypes in Southern Croatian Male Population…) с 166 хорватскими, а в YHRD представлены 1320 хорватских, 1270 польских, 1377 российских, 1774 германских, по 200 словацких и русинских (фактически – закарпатско-украинских) с территории Сербии, по 100 албанских и македонских, 191 греческий, 154 румынских, а недавно добавились 154 западноукраинских из Львовской области и 486 австрийских. Для полноты картины стоит привлечь 1262 итальянских и 194 литовских. По Беларуси у меня есть данные по 1097 коренным уроженцам из базы Центра судебной медицины. Всего около 10 тысяч — это уже серьезный массив данных! Правда, ни один из этих гаплотипов не тестировался на L147.2 и даже на L621, равно как и на L260. По счастью, карпийские гаплотипы на 17 локусах довольно четко отделяются от других гаплогрупп (погрешность в их выявлении я оцениваю на уровне порядка 1-2%, главным образом за счет пресловутого кластера Disles). Гаплотипы западнославянского кластера R1a1 отделяются от общеславянского М458+L260- чуть менее надежно, но все же с приемлемой точностью.сего в указанных источниках мне удалось выявить предположительно 1363 карпийских 17-локусных гаплотипов. Еще несколько остаются под вопросом. В частности, в Хорватии и Сербии выявлены 3 гаплотипа, профиль которых из всех известных мне модальных ближе всего к карпийскому, за исключением локуса Gata-H4, в котором вместо 11 повторов, характерных для I2a1b, имеется 9. Не имея снипов, трудно сказать, идет ли речь о специфической мутации в карпийской ветви, или о совершенно иной гаплогруппе. Я решил их не учитывать.Если интересны суммарные результаты, то они такие (в порядке убывания частоты):n N % популяция65 179 36,3 Сербия497 1486 33,4 Хорватия120 404 29,7 Черногория34 154 22,1 Украина (Львов)151 693 21,8 Беларусь (центр — юг)33 154 21,4 Румыния21 100 21,0 Македония38 200 19,0 Русины из Сербии (Нови Сад)29 200 14,5 Словаки из Сербии (Нови Сад)21 191 11,0 Греция (север)40 405 9,9 Беларусь (запад — север)9 100 9,0 Албания121 1377 8,8 Россия109 1270 8,6 Польша24 486 4,9 Австрия6 194 3,1 Литва37 1774 2,1 Германия4 1262 0,3 Италия1359 10629 Как решается вопрос о генетическом разнообразии по моей методике? Если рассмотреть наиболее удаленные гаплотипы, то, даже отбразывая несколько сомнительных на удалении в 14-15 “калиброванных” шагов, 2 гаплотипа (из свердловской выборки русских и из южной Хорватии) удалены от модального более чем на 10 шагов. На удалении от 9 до 10 шагов находятся 9 гаплотипов (4 хорватских, 1 сербский, 1 македонский, 1 русинский, 1 свердловский и 1 белорусский). В интервал от 8 до 9 шагов попадают 12 гаплотипов (4 хорватских, 2 черногорских, 1 сербский, по 1 словацкому, германскому, австрийскому, польскому, свердловскому), от 7 до 8 шагов – 30 гаплотипов (из них 14 хорватских и 2 черногорских, а белорусских всего 3). В целом среди наиболее удаленных гаплотипов доля балканских примерно такая же, как и во всей выборке (55% против 56), тогда как доля белорусско-украинско-словацких – явно ниже (15% среди удаленных и 21 – во всей выборке). При всех неточностях и условностях такой методики, говорить о более высоком разнообразии гаплотипов в ареале пражской культуры по сравнению с Балканами нет никаких оснований. Скорее – наоборот.Теперь по поводу оценки возраста. Самым надежным репером является примерная синхронность с западнославянским кластером R1a1-L260. Эта синхронность подтверждается и по выборке из YHRD (вкупе с белорусскими и некоторыми другими данными). Предположительно к этому кластеру можно отнести чуть более 300 гаплотипов, из которых 166 – польских и 33 – германских. Наиболее удаленные 8 гаплотипов попадают в тот же интервал, что и в карпийском кластере: от 7.3 до 10.3 шагов.Распределение западнославянского кластера позволяет привязать его возраст к событиям, фиксируемым археологически. Верхняя граница – это формирование суковско-дзедзицкой культуры (некоторые считают ее частью пражской), т.е. 1500 — 1400 лет назад. Но, учитывая возможное наличие этого кластера в России и Хорватии, скорее – раньше. Нижняя граница вряд ли может быть древнее формирования лужицкой культуры, т. е. 3600 – 3000 лет назад. Думаю, что в нижнюю половину этого интервала укладывается и возраст карпийского кластера. Я бы оценил его в 3000 лет плюс-минус пятьсот. Вокруг Карпат — это время протофракийской каннелированной и штампованной керамики, культурных общностей гава-голиграды и белогрудовка-чернолесье. На западе Балкан – культура глазинац, мимо которой прокатывается 3,3 – 3,2 тыс лет назад волна будущих «народов моря» (чака – медиана). Зарождение карпийского кластера можно связывать с любой из них – практически на равных основаниях. Учитывая господство в это время обряда трупосожжения, ископаемая ДНК тут вряд ли поможет.Тем более от рассуждений о более ранней истории I2a1, равно как и от комментариев по поводу Ваших предположений на этот счет, предпочту воздержаться.
Почему Вы оперируете только 17 маркернымі гаплотипами, когда известны 37, 64,111 маркерные гаплотипы I2a1b3?Это не есть математическое доказательство. Предлагаю Вам повторить Ваше упражение с 1363 17-локусными гаплотипами — но на этот раз Вы должны расчитать интер — и интракладовые дисперсии «карпийских» гаплотипов по странам. Тогда мы сравним с моей статистикой дисперсии 37-64 локусных гаплотипов. Рекомендую использовать для этих целей .
Вячеслав Носевич:
Насчет «Арлекина» — если честно, то мне лень возиться. Достаточно уже повозился. пока вытаскивал гаплотипы из YHRD (сервис там еще тот…) Если хотите — пришлю все 1363 Вам, а Вы считайте как хотите и чем хотите. Только мне потом расскажите, что получилось.
Комментарий Вячеслава Малиновского:
«Если бы у нас были надежные выборки, порядка сотен длинных гаплотипов в каждой ветви, тестированных по всем Y-снипам, мы увидели бы, что САМЫЕ УДАЛЕННЫЕ ОТ МОДАЛЬНОГО различаются тем сильнее, чем древнее кластер. Проблема лишь в том, что такие выборки появятся очень нескоро.» Если вы еще раз посмотрите что же писал Вадим, то увидите, что речь шла именно о выборках из сотен длинных гаплотипов, на основании которых и строились филогенетические деревья.


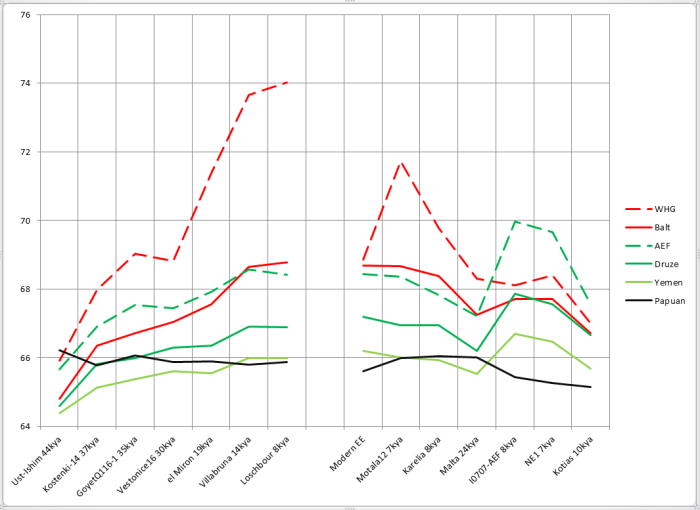



 При сравнении видно, что связь с восточной (и в первую очередь Юго-Восточной) Азией несколько ослабла, а связь с западными выборками (как европейскими, так и ближневосточными) слегка усилилась. Однако разница невелика и из-за этого сравнительная карта выглядит некрасиво. Чтобы избежать загромождения излишними иллюстрациями, ее не привожу.
При сравнении видно, что связь с восточной (и в первую очередь Юго-Восточной) Азией несколько ослабла, а связь с западными выборками (как европейскими, так и ближневосточными) слегка усилилась. Однако разница невелика и из-за этого сравнительная карта выглядит некрасиво. Чтобы избежать загромождения излишними иллюстрациями, ее не привожу.

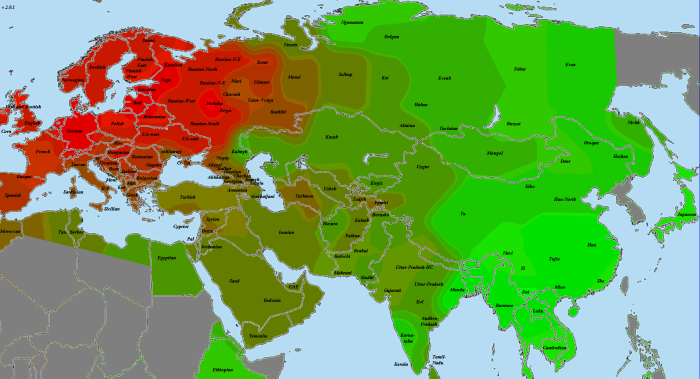 Как видите, прибытие Villabruna никакого переворота не произвело. Как и раньше, с ходом времени сходство с циркумбалтийцами усиливалось, с восточноазиатами — ослабевало, с ближневосточниками — то ли слегка усиливалось, то ли медленно ослабевало, но медленнее, чем с восточноазиатами.
Как видите, прибытие Villabruna никакого переворота не произвело. Как и раньше, с ходом времени сходство с циркумбалтийцами усиливалось, с восточноазиатами — ослабевало, с ближневосточниками — то ли слегка усиливалось, то ли медленно ослабевало, но медленнее, чем с восточноазиатами.


























Для отправки комментария необходимо войти на сайт.